研究内容
私たちの生活する環境には知らず知らず刺激性化学物質がたくさん存在する

私たちは、日々の生活の中で何気なく多くのストレスにさらされていますが、ほとんどの場合何事もなく生活ができていると思います。しかし、知らず知らずのうちに、農薬や、植物の花粉などの異物、カビなどの胞子類、PM2.5に挙げられる大気汚染物質、タバコの煙、車の排気ガス、紫外線など様々な物質にさらされています。あげればキリがないですが、こういうものがある環境においても、私たちは健康で、文化的な生活を送ることができています。一方で、これらの感知機構が崩れるとどうなるでしょうか?例えば、紫外線は主にゲノムDNAに損傷を与えますが、この認識機構が破綻すると、重篤な光過敏症が起きます。これらは先天性の遺伝病としても知られますが、それほど私たちの環境中にはストレスが多いということを認識していただければと思います。
環境を感知して、応答する分子メカニズム:環境転写因子って
一方で、何もストレスがない時、オオカミ少年のように、「大変だ!」と体が応答した場合はどうなるでしょうか?想像しやすい現象としてアレルギーがあります。普通は、ストレスにならない、ピーナッツ等によって、重篤な反応が引き起こされてしまいます。こういう現象が、何もしていなくても発現することとなれば、もはやお薬なしには通常の生活が送れなくなります。中には、ただ服を着ただけで、その刺激が針の様に感じるような病気もあります。
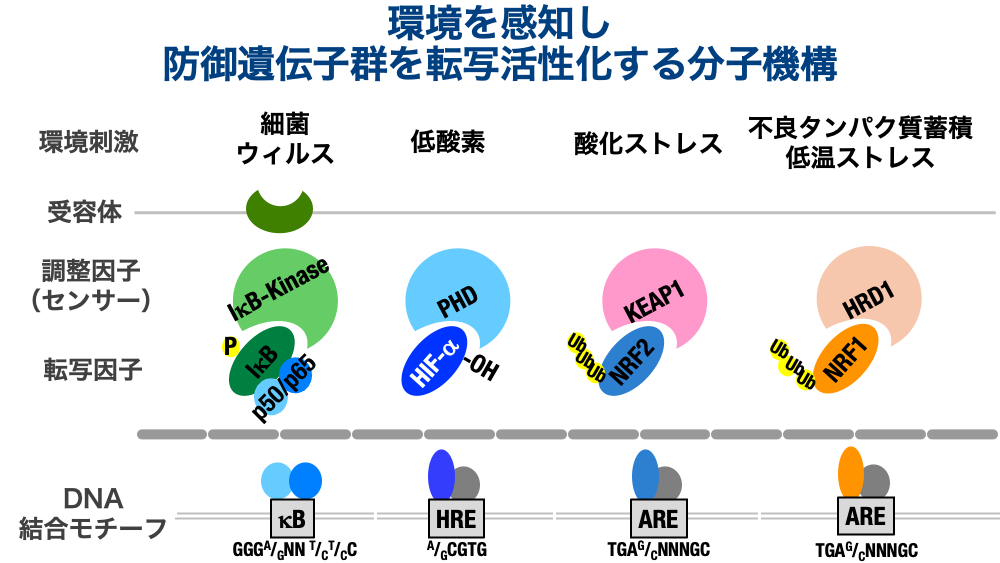
私どもは、物理的な刺激よりも、化学物質を感知して、応答する生体のしくみについて研究をしております。主に、細菌やウイルスの感染を認識する自然免疫機構、酸素濃度の低下等を認識するPHD-HIF系、酸化ストレス物質や、タンパク質やDNAに対して求核的に作用するような化合物を認識するKEAP1-NRF2系、不良タンパク質蓄積や低温ストレスを認識するNRF1などを研究対象としています。これらの転写因子は、刺激がない状態においては、常にユビキチンプロテアソーム系で分解制御を受けているとされ、タンパク質の存在量は非常に低く保たれています。すなわち、刺激がない状態にておいては、適応反応を司どる防御タンパク質は転写誘導されません。
しかし、ひとたび、センサータンパク質が刺激を感知すると、分解制御されていたこれらの環境転写因子は、ユビキチン修飾が低下することで、分解制御を免れます。これらのタンパク質には核移行シグナル配列が存在するため、速やかに核内へ移行して、特定の遺伝子配列に結合して、標的遺伝子の転写を活性化します。
注意:私たちがここで研究対象としているのは、NRF1:Nuclear factor erythroid-derived 2-related factor 1、およびNFE2L1: Nuclear factor erythroid-derived 2 like 1となります。時にNuclaer Resperatory Factor と勘違いされることがありますので・・・
環境応答ストレスセンサーNRFファミリー
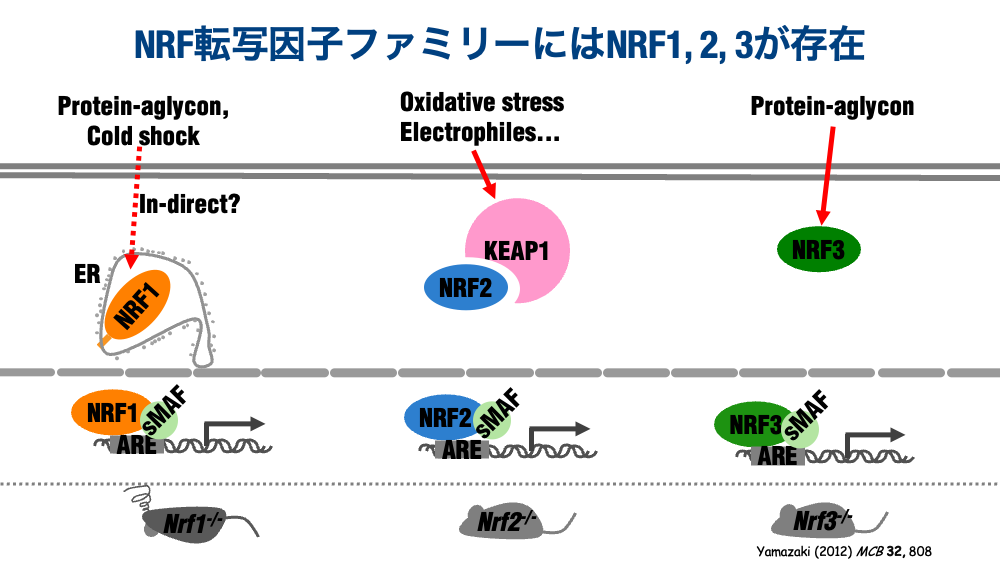
先ほど、NRF1とNRF2について少し触れましたが、ののNRF転写因子ファミリーは脊椎動物で1から3まで存在します。NRF1が不良タンパク質や低温ストレス、NRF2が酸化ストレスや、タンパク質やDNAに対して求核的に作用するような化合物、NRF3が不良タンパク質を認識して、個体や細胞に抵抗生を付与する、防御タンパク質を統一的に誘導することが明らかとなっています。
これらの転写因子の遺伝子欠失マウスは2000年代に相次いで作出されましたが、NRF2とNRF3の全身欠失マウスは、生育にも生殖にも影響が全く見られない傍ら、NRF1のみが出生後すぐに死亡してしまうので、長らく研究が遅れていました。
NRF1は標的遺伝子を抑制的に制御する???
長い間、NRF1は抗酸化タンパク質を発現誘導するNRF2と同様に、外部からの刺激によって安定化して、これまたNRF2と同様に抗酸化タンパク質を発現誘導すると考えられてきました。NRF1はNRF2と同様、抗酸化応答配列(ARE, antioxidant response element or EpRE, electrophile response element)(TGA[G/C]NNNGC)に結合することが&deco(i,,,){in vitro};のEMSA解析で明確になっていたからです。しかし、NRF1の全身遺伝子欠失マウスは出生後すぐに死亡してしまうのに対して、NRF2は普通に生まれてくるばかりか、生殖能力も有すると表現系が全く違います。
当時存在しなかった、NRF1タンパク質を認識する抗体を樹立したところ、どうやら、マウスの肝においては、通常状態でもNRF1タンパク質が十分存在していることが明らかになりました。すなわちNRF1は未刺激状態でも何らかの転写調節に関与すると思われました。現在では、市販の抗体でNRF1タンパク質を検出することができます。
個体におけるNRF1の機能解析を可能にしたCYP1A1-Creシステム
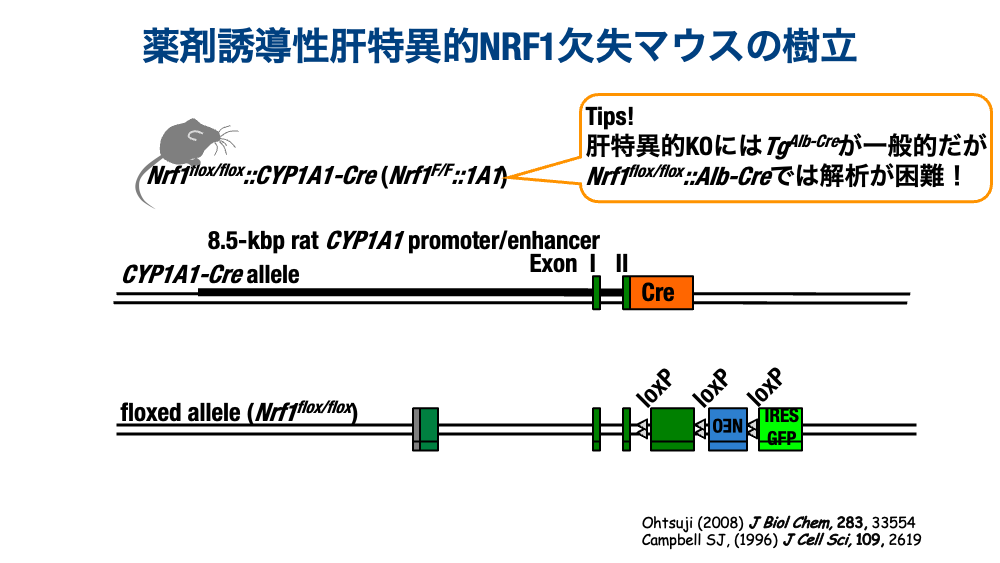
個体におけるNRF1の研究のために、早期に条件付き遺伝子欠失システムを活用してNRF1の遺伝子欠失が試みられました。当初、肝特異的にCreリコンビナーゼを発現するAlbmin-Creトランスジェニックマウス、神経系特異的にCreリコンビナーゼを発現するNestin-Creトランスジェニックマウスとの掛け合わせで解析が進みました。私どもは肝に着目した解析をしておりましたので、Nrf1 F/F::Albmin-Creマウスで当初実験を進めていましたが、表現系が安定しないことに気がつきました(ただ単に、怠け者だから・・・?)そこで、大胆にCreのシステムを変更して、出生後にNRF1を欠失できるマウスを作出していきました。掛け合わせたのはMx-Cre、Tamoxifenで誘導されるCreERシステムなどを試しています。それぞれに問題があって、最終的に採用したのはCYP1A1-Creというシステムでした。このシステムは、CYP1A1を誘導するAhRレセプターと結合する、3-Methylcholanthrene(3-MC)を皮下に注射することで、出生後の成獣で肝特異的に遺伝子を欠失できるシステムです。任意の週齢で3-MCを注射し、2週間待つことで、NRF1が十分欠失することを明らかにしています。
このユニークな薬剤誘導的肝特異的NRF1欠失マウスの作出によって、以前のAlbumin-Creマウスの解析に比べて、顕著な3つの異常を見出しました。
・顕著な脂肪蓄積:Albmin-Creマウスにおいても、著名な脂肪肝が観察されていたが、その脂肪の組成などについては明確になっていませんでした。私どもは、質量分析器を用いて、脂質の素性を明らかにし、その中で、FADS3がNRF1によって制御されることを見出しています。
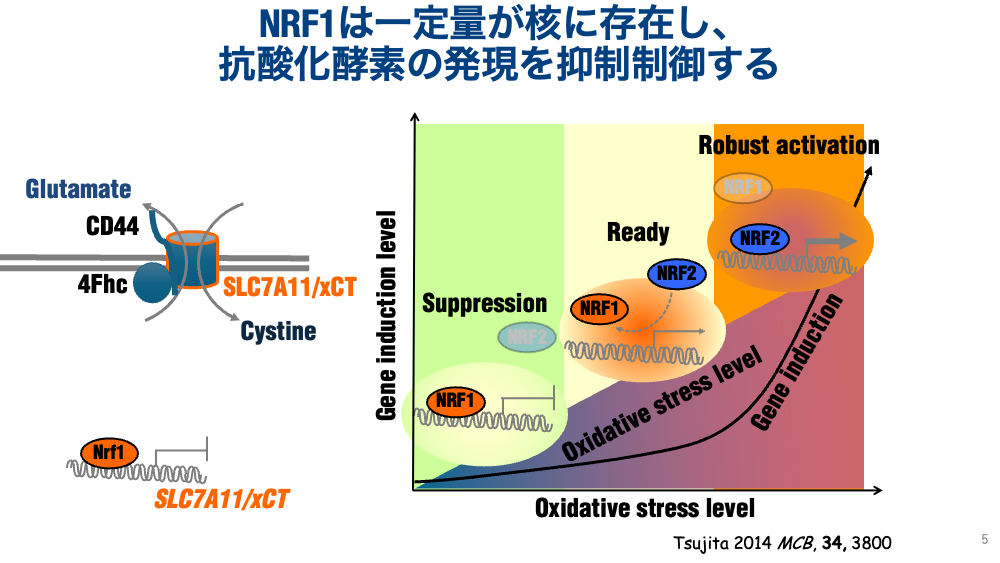 ・抗酸化物質グルタチオンが蓄積する?この原因は、xCT(細胞外からシスチンを取り込む受容体)のectopicな発現上昇にあることを突き止めました。つまり、NRF1が積極的にxCTを抑制制御していたわけです。xCTはがんマーカーとして用いられていますから、NRF1の発現変動も病変に関係するのではないかと思われています。私たちは、NRF1が通常、抗酸化酵素タンパク質を抑制制御することを提唱しており、積極的な抑制制御こそが、外来ストレッサーの認識よりも大事ではないかと考えています。例えるなら、ブレーキが壊れた車と、アクセルが壊れた車どっちが危険ですかということです。アクセルが壊れていれば、車は走りませんので、ただ乗るだけですが、ブレーキが壊れた車は、かなりの確率で、事故に遭います。どっちが大事かはわかると思います。
・抗酸化物質グルタチオンが蓄積する?この原因は、xCT(細胞外からシスチンを取り込む受容体)のectopicな発現上昇にあることを突き止めました。つまり、NRF1が積極的にxCTを抑制制御していたわけです。xCTはがんマーカーとして用いられていますから、NRF1の発現変動も病変に関係するのではないかと思われています。私たちは、NRF1が通常、抗酸化酵素タンパク質を抑制制御することを提唱しており、積極的な抑制制御こそが、外来ストレッサーの認識よりも大事ではないかと考えています。例えるなら、ブレーキが壊れた車と、アクセルが壊れた車どっちが危険ですかということです。アクセルが壊れていれば、車は走りませんので、ただ乗るだけですが、ブレーキが壊れた車は、かなりの確率で、事故に遭います。どっちが大事かはわかると思います。
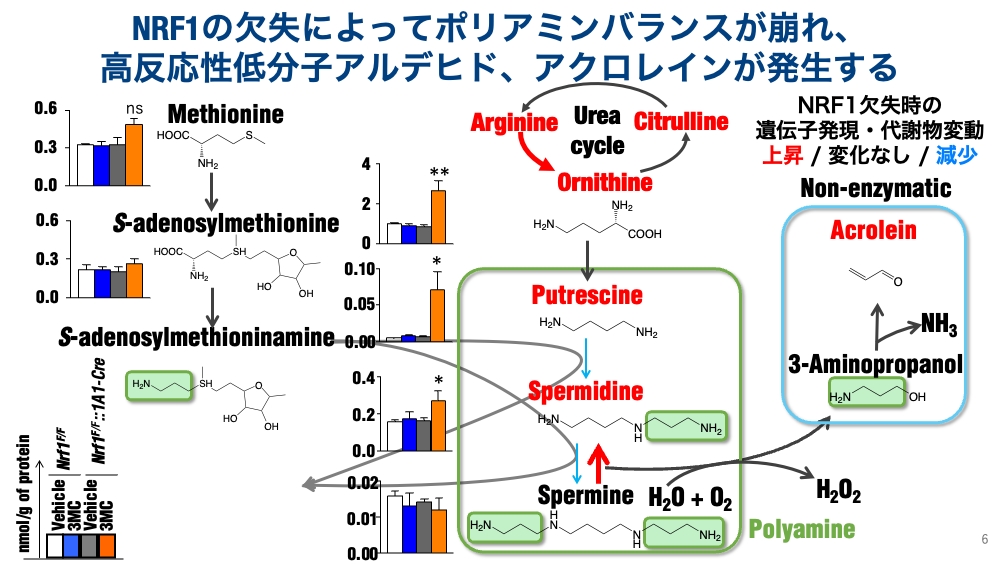 ・ポリアミン代謝の異常。低分子代謝物の解析から、ポリアミンの代謝バランスに異常があることも見出しました。ポリアミンはプトレッシン、スペルミジン、スペルミンの三種が脊椎動物には含まれますが、最も長鎖のスペルミンが多く分解されていることがわかりました。この分解にはスペルミンオキシダーゼ(SMOX)とポリアミンオキシダーゼ(PAOX)によって分解される、2経路が存在するのですが、NRF1の欠失時にはSMOX経路の方が亢進する様子が明らかになりました。このSMOXの亢進は組織的には大きな問題を孕んでおり、スペルミンの分解によって、過酸化水素、アンモニア、そして非酵素的にアクロレインという不飽和アルデヒドが出てきます。このアクロレインが細胞内因的に蓄積することが、NRF1欠失による肝障害の原因になることまでを突き止めています。
・ポリアミン代謝の異常。低分子代謝物の解析から、ポリアミンの代謝バランスに異常があることも見出しました。ポリアミンはプトレッシン、スペルミジン、スペルミンの三種が脊椎動物には含まれますが、最も長鎖のスペルミンが多く分解されていることがわかりました。この分解にはスペルミンオキシダーゼ(SMOX)とポリアミンオキシダーゼ(PAOX)によって分解される、2経路が存在するのですが、NRF1の欠失時にはSMOX経路の方が亢進する様子が明らかになりました。このSMOXの亢進は組織的には大きな問題を孕んでおり、スペルミンの分解によって、過酸化水素、アンモニア、そして非酵素的にアクロレインという不飽和アルデヒドが出てきます。このアクロレインが細胞内因的に蓄積することが、NRF1欠失による肝障害の原因になることまでを突き止めています。
NRF1を制御する物質はなんだ?
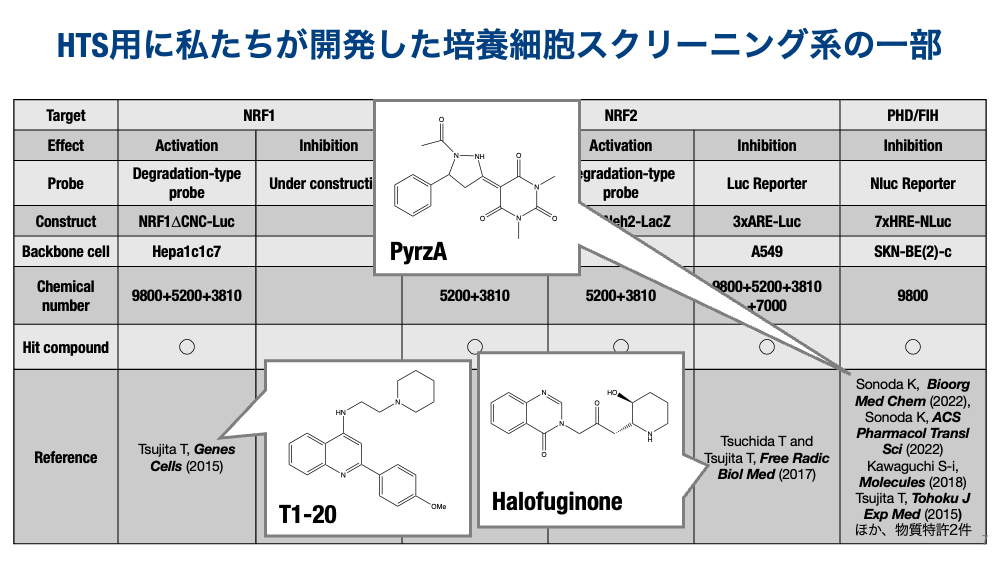
NRF2の研究は爆発的に進展する一方、NRF1の研究は、プロテアソームの構成因子を統一的に誘導できることが明らかとなって、一気にセンセーショナルな転写因子になりましたが、やはり制御が複雑なので、なかなか解析が進まないのが現状です。その原因は、NRF1を特異的に誘導する物質が不明であることにあると考えました。私たちは、NRF1の安定化を高感度に検出できるレポーター細胞を樹立して、数万化合物から特異的な活性化物質(T1−20)を取得しました。現在は、T1-20以外のヒット化合物の精査や、標的タンパク質の同定を進めています。
NRF1を制御する物質をもっと欲しい!
NRF1を制御できる物質は、食品などにも含まれる可能性があります。そこで、農産物等の評価を進めています。一連の機能性評価を通して、地産商品のブランディングに貢献できると考えています。将来は、取得した機能性成分を活用して、創薬分野のみならず、健康食品や化粧品への応用研究も進め、農学分野から健康長寿社会へ資する研究を推進したいと考えています。
抗酸化タンパク質は多ければいいか?
抗酸化タンパク質を多く保持することは、お酒に酔いにくくなったり、がんになりにくくなったり良いことばかりと考えられていますが、ある種のがんはそれを逆手にとって、抗がん剤に対して抵抗性を獲得しています。抗酸化タンパク質は刺激に応じて、適切な時、適切な臓器・場所で誘導され、悪者を処理したら速やかに消える必要があるわけです。
評価方法について
これまでは、遺伝子改変マウスを用いた解析が主体でしたが、もっと簡便に評価をしたいので、小型魚類を用いた実験系を導入しております。私も魚好きが高じて、小型魚類を導入しましたが、興味がある方、共同研究等いつでもお待ちしております。
研究設備

日々実験する実験台です。ここは、タンパク質の電気泳動装置と膜への転写装置があります

大腸菌を無菌培養するためのクリーンベンチです。研究室の中に置いてあり、遺伝子の機能解析に必要なプラスミド構築を大腸菌に作ってもらうために必須です。ただ、ガスバーナーの下で実験しても大腸菌は無菌的に扱うことができますけど・・・

遺伝子組換え大腸菌を培養するための、いわば大腸菌のための家です。37ºCで設定されてあります。場合によっては、20ºCなどの培養も実施します

辻田忠研では、通常のPCRを4台、定量PCRを2台おいています。定量PCRは増幅した遺伝子断片を蛍光色素で検出し、その度合いによって、遺伝子増幅を高感度で検出する機械です。刺激によって、遺伝子が誘導されて、発現変動があるのかないのかを検出するのに便利です。もっとも、最終確認はタンパク質量で検証するのですが・・・

遺伝子の増幅のために、古いのから新しいものまでPCRを4台保有しています。長い断片など、さくっと増えないときには、DNA合成酵素を変更するのも一つの手ですが、PCRを変えてみるのも効く場合があります。教室では、それぞれに推しPCRがあるとか、ないとか。こちらは古い方のPCRさんですね。

DNAを増幅したあと、実際に増幅しているかをアガロースで電気泳動で分離して、染色したDNAを確認するための装置です。基本的には紫外線を使用して蛍光シグナルとして確認するのですが、最近では、紫外線がDNAに対してダメージを多く与えるので、緑色の光を当てて、DNAに優しく撮影する方法もとっています。ただ、この緑色の光、すこぶる眩しいです。

佐賀大学農学部では、研究室の蛇口まで(おいしい?)純水が供給されています。さらに精製した水を作り、分子生物学実験で使用できるグレードの水を調整する機械です。飲むとお腹を壊すという都市伝説があります。

こちらは培養細胞を無菌的に培養するための、セーフティーキャビネットです。P2レベルの感染性生物までを使用できます。辻田忠研では、レンチウイルスやレトロウイルス遺伝子導入の実験まで実施できます!

培養細胞は、それ自体が透明ですので特殊な光を当てて、観察する必要があります。こちらの顕微鏡は、位相差観察をする顕微鏡です。デジタルカメラが後ろに内蔵されていて、覗かなくても、細胞をモニターで観察できます。私は、のぞく派なので、必ず目視で確認をします。

研究費を獲得して、そのほとんどを注ぎ込んで購入した、倒立蛍光顕微鏡です。こちらは、こだわって、XYZの制御が全て電動になっています。すなわち、大きなサンプルもタイリングすることで高精細な画像を取得できます。また、計時的に自動で写真を撮影することもできますので、タイムラプス観察なども可能です。なお、明視野観察は、位相差観察と微分干渉観察を行うことができます。これらと、DAPI、FITC、TRITCフィルターも装備していますので、最大4枚の写真を撮影できます。もちろん予め、観察する細胞の座標を指定しておけば、指定された時間に、その細胞だけを、必要な蛍光染色で撮影できます。辻田研ビジュアル系を支えています。

基本的には、培養細胞への遺伝子導入はリポフェエクションを利用して導入していますが、効率よく導入したい場合には、エレクトロぽレーションを活用します。こちらはピペットタイプのエレクトロポレーションでして、簡便に遺伝子を導入できます。ただ、チップが金なのか何なのか、高いです。

佐賀大に来て、清水の舞台かr飛び降りる勢いで購入したプレートリーダーです。任意の波長での吸光度・蛍光測定、発光測定が可能です。また、96穴に播種した細胞の明視野、FITC、TRITC観察画像を取得することができます。スクリーニングに大活躍で、共通機器へ供出していますけど、大人気です。

培養細胞は基本的に、実験室のクリーンベンチで取り扱います。すなわち酸素濃度は21%になっています。私たちの体内は5-7%と低酸素状態にあるので、生体内の現象を見るために、時折、低酸素で細胞培養を行う必要があります。私どもが低酸素環境で培養細胞の世話をすると、確実に私たちが死にます。それを避けるために、低酸素で無菌培養をできる培養チャンバーを導入しています。こちらのチャンバーは酸素濃度を5-21%の間で制御可能です。なお、二酸化炭素濃度については、5%で運用しています。もちろん、庫内温度は37ºCです。

こちら、スキャナー型の顕微鏡で、プレートに播種した培養細胞を計時的に観察することができます。特に、化合物等をかけた時、いつ、どのようにして死んでいくのかを観察するときに重宝します。先ほどの、低酸素チャンバーの中に設置して、細胞の観察に使用しています。最近では辻田研You Tuberができるとか何とか

こちらは、一般の培養細胞を培養するインキュベーターです。庫内は、湿度100%、CO2濃度5%、温度37ºCに保たれています。下には、同様のインキュベーターがありますが、酸素濃度のコントロールが可能なものです。こちらでは1.0-20%の酸素濃度をコントロールすることができます。インキュベーターの壁に手袋が引っかかっていますが、辻田研では手袋を破れるまで使ってもらいますので、個人の名前が書いてあります。使えるものは最後まで使いましょう!

辻田研では多くの冷凍庫があります。こちらは、-30度でサンプルや、試薬保管する冷凍庫です。部屋のあちこちに冷凍庫があります。個人個人に引き出しが一つ与えられ、実験サンプルを保存してもらっています。

こちらは、-80度で保管するディープフリーザーです。基本的にRNAやタンパク質、培養細胞、臓器などを保管しています。