Research
Irritant chemicals surround us in our daily lives — without us even knowing

We are constantly exposed to a wide variety of stressors in our daily lives, yet in most cases we go about our days without any obvious ill effects. Unbeknownst to us, however, we are continuously exposed to substances such as agricultural pesticides, foreign particles from plant pollen, mold spores, atmospheric pollutants including PM2.5, cigarette smoke, vehicle exhaust fumes, and ultraviolet radiation — the list is endless. Despite this, we are able to maintain good health and lead culturally rich lives. But what happens when these sensing mechanisms break down? Ultraviolet light, for example, primarily damages genomic DNA, and when the recognition system that detects this damage fails, severe photosensitivity disorders can result. These are also known as congenital genetic diseases, and this illustrates just how many environmental stressors surround us.
Molecular mechanisms of environmental sensing and response: What are "environmental transcription factors"?
On the other hand, what happens when the body responds with alarm — like the boy who cried wolf — even when there is no real stress? A familiar example is allergy: peanuts or other substances that pose no danger to most people can trigger severe reactions in sensitized individuals. If such responses occur spontaneously without any external trigger, normal daily life becomes impossible without medication. There are even diseases in which the simple act of wearing clothes feels like needles against the skin.
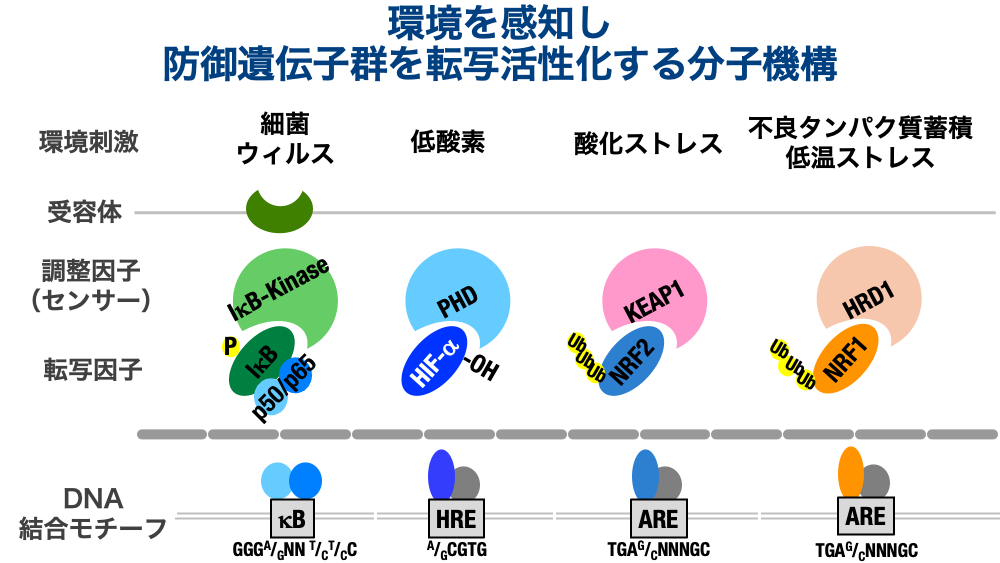
Our laboratory studies the biological mechanisms by which organisms sense and respond to chemical substances, rather than to physical stimuli. Our main research subjects include: the innate immune system, which detects bacterial and viral infections; the PHD–HIF axis, which senses decreases in oxygen concentration; the KEAP1–NRF2 system, which recognizes oxidative stressors and compounds that act nucleophilically on proteins and DNA; and NRF1, which responds to misfolded protein accumulation and cold stress. These transcription factors are continuously degraded by the ubiquitin–proteasome system in the absence of a stimulus, keeping their protein levels extremely low. In other words, when no stimulus is present, the defensive proteins that orchestrate adaptive responses are not transcriptionally induced.
However, once a sensor protein detects a stimulus, the ubiquitin modification of these environmental transcription factors decreases, allowing them to escape degradation. Because these proteins contain nuclear localization signals, they rapidly translocate to the nucleus, bind to specific gene sequences, and activate transcription of their target genes.
Note: The subjects of our research here are NRF1 (Nuclear factor erythroid-derived 2-related factor 1) and NFE2L1 (Nuclear factor erythroid-derived 2 like 1). They are sometimes confused with Nuclear Respiratory Factor, so please be aware of the distinction.
The NRF Family: Stress Sensors for Environmental Response
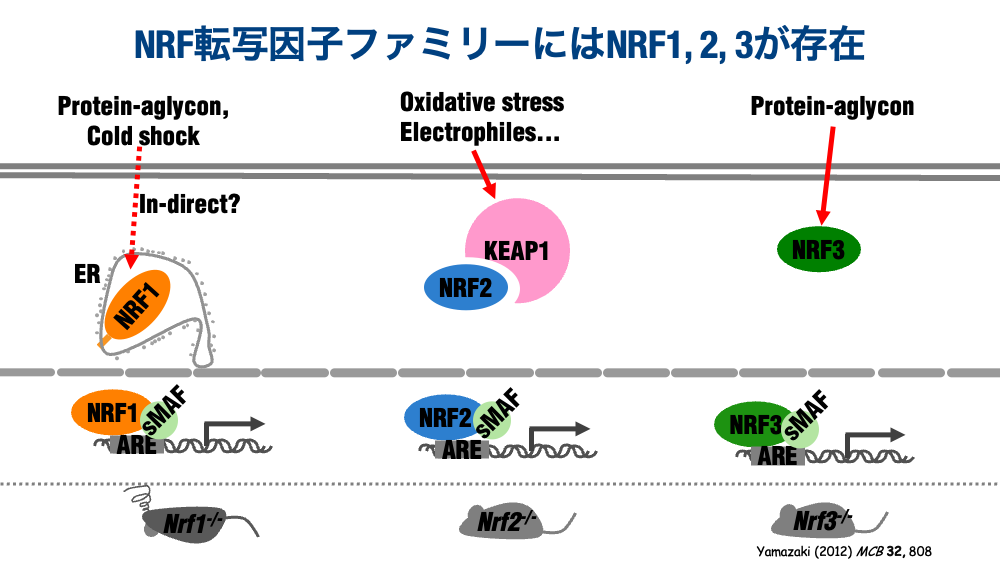
As mentioned above, the NRF family of transcription factors comprises three members (NRF1–3) in vertebrates. It has been established that NRF1 responds to misfolded proteins and cold stress, NRF2 to oxidative stress and compounds acting nucleophilically on proteins and DNA, and NRF3 to misfolded proteins — each coordinately inducing a set of cytoprotective genes that confer resistance to cells and organisms.
Whole-body knockout mice for each of these transcription factors were generated in succession during the 2000s. While mice lacking NRF2 or NRF3 show no impact on growth or reproduction, NRF1-null mice die shortly after birth — a finding that long delayed progress in NRF1 research.
Does NRF1 Repress Target Gene Expression?
For a long time, NRF1 was thought — like NRF2, which induces the expression of antioxidant proteins in response to external stimuli — to be stabilized by stimuli and to similarly induce antioxidant protein expression. This assumption was based on clear in vitro EMSA evidence showing that NRF1, like NRF2, binds the antioxidant response element (ARE, also called EpRE; TGA[G/C]NNNGC). However, while whole-body NRF1-null mice die immediately after birth, NRF2-null mice are born normally and are even fertile — a completely different phenotype.
When we generated an antibody recognizing the NRF1 protein — something that did not exist at the time — we found that NRF1 protein is present in mouse liver even under unstimulated conditions. This suggested that NRF1 is involved in transcriptional regulation even without stimulation. Today, commercially available antibodies can be used to detect NRF1 protein.
The CYP1A1-Cre System: Enabling In Vivo Functional Analysis of NRF1
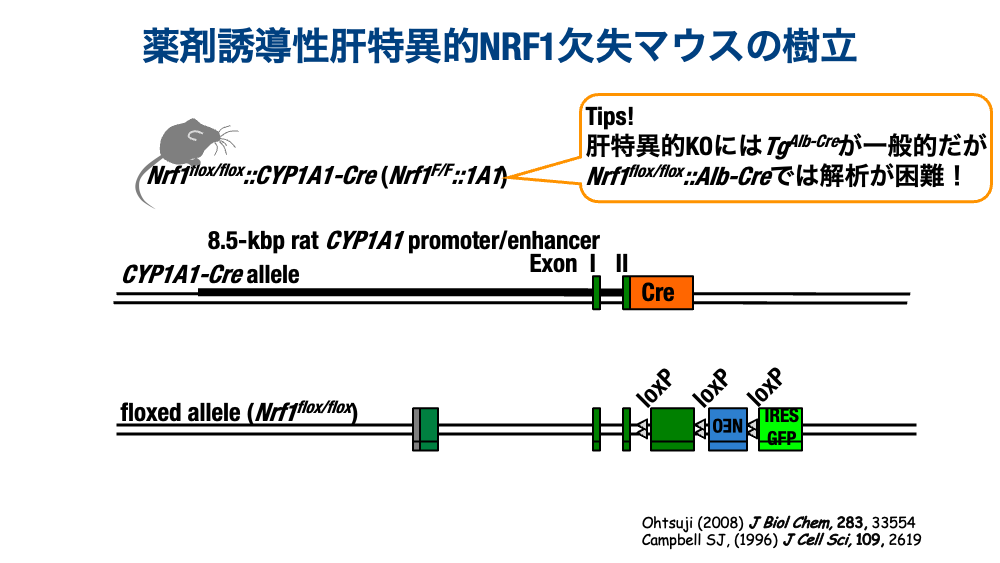
To study NRF1 function in vivo, conditional gene knockout systems were employed early on. Initial studies were conducted by crossing NRF1-floxed mice with Albumin-Cre (liver-specific) and Nestin-Cre (nervous system-specific) transgenic mice. Focusing on the liver, we initially used Nrf1 F/F::Albumin-Cre mice, but soon noticed that the phenotype was inconsistent (perhaps simply because the mice were lazy…?). We therefore boldly switched Cre systems to generate mice capable of postnatal NRF1 deletion. We tested Mx-Cre and tamoxifen-inducible CreER systems, each of which had its own drawbacks. The system we ultimately adopted was CYP1A1-Cre, in which subcutaneous injection of 3-methylcholanthrene (3-MC) — a ligand of the AhR receptor that induces CYP1A1 — achieves liver-specific gene deletion in adult mice at any chosen age. We confirmed that NRF1 is sufficiently deleted two weeks after 3-MC injection.
Using this unique drug-inducible, liver-specific NRF1-deficient mouse model, we identified three prominent abnormalities compared to analyses with Albumin-Cre mice:
- Marked fat accumulation: Striking hepatic steatosis had also been observed in Albumin-Cre mice, but the composition of the accumulated lipids had remained unclear. We characterized the lipids by mass spectrometry and found that FADS3 is regulated by NRF1.
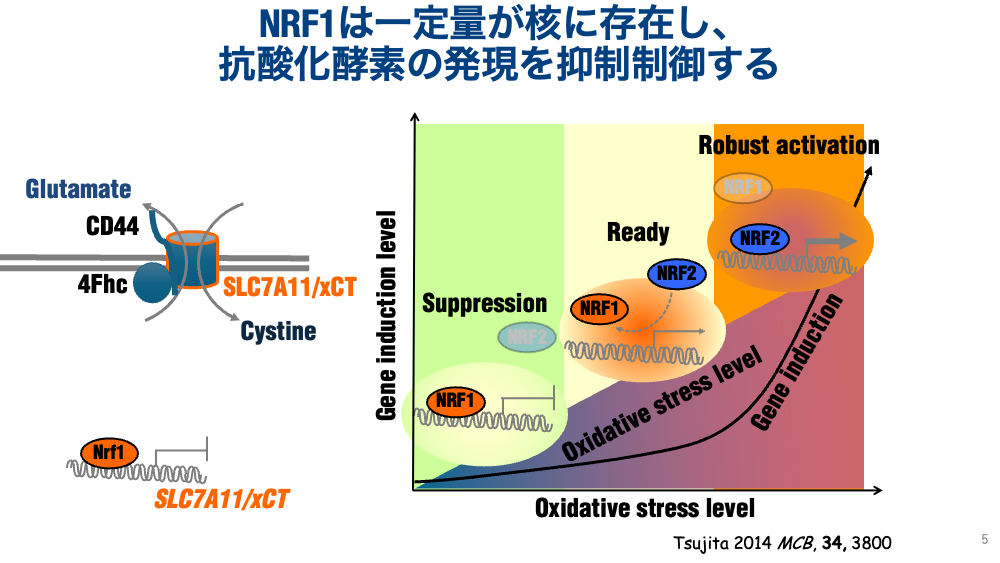
- Accumulation of the antioxidant glutathione: We traced this to ectopic upregulation of xCT (a receptor mediating cystine uptake from outside the cell), demonstrating that NRF1 normally suppresses xCT. Because xCT is used as a cancer marker, changes in NRF1 expression may be linked to pathological lesions. We propose that NRF1 actively represses antioxidant enzyme proteins, and that this active repression is more important than recognition of exogenous stressors — analogous to asking whether a car with broken brakes or a car with a stuck accelerator is more dangerous. A car with a stuck accelerator simply won't move, whereas a car with no brakes will very likely cause an accident.
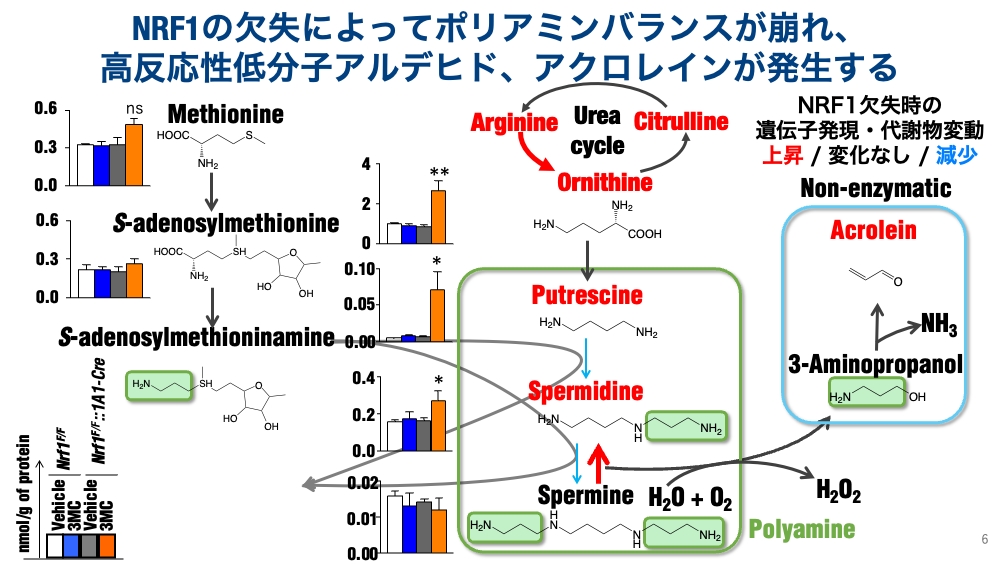
- Dysregulation of polyamine metabolism: Analysis of low-molecular-weight metabolites revealed an imbalance in polyamine metabolism. Vertebrates contain three polyamines — putrescine, spermidine, and spermine — and we found that spermine, the longest-chain polyamine, was most extensively degraded. Spermine degradation proceeds via two pathways, catalyzed by spermine oxidase (SMOX) and polyamine oxidase (PAOX); upon NRF1 deletion, the SMOX pathway appears to be preferentially activated. This SMOX upregulation is highly problematic at the tissue level: spermine degradation produces hydrogen peroxide, ammonia, and — non-enzymatically — acrolein, an unsaturated aldehyde. We have identified the intracellular accumulation of acrolein as a cause of the liver injury observed upon NRF1 deletion.
What Substances Regulate NRF1?
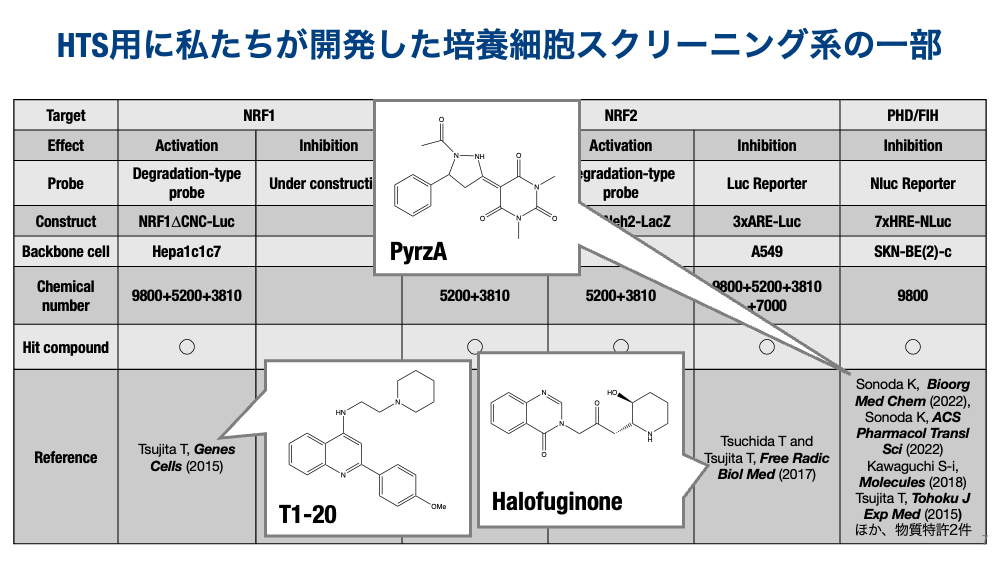
While research on NRF2 has expanded explosively, NRF1 research has become more prominent since it was shown to coordinately induce proteasome subunit genes — making it a suddenly high-profile transcription factor — but the complexity of its regulation continues to impede progress. We believed this was largely because the substance that specifically induces NRF1 remained unknown. We therefore established reporter cell lines capable of detecting NRF1 stabilization with high sensitivity, and obtained specific activators (T1-20) from screening tens of thousands of compounds. We are currently working on further characterization of hit compounds other than T1-20 and on identifying their target proteins.
We want more substances that regulate NRF1!
Substances capable of regulating NRF1 may be present in foods and agricultural products. We are therefore proceeding with functional evaluation of agricultural products, particularly those from Saga Prefecture. Through these evaluations, we believe we can contribute to the branding of locally produced goods. In the future, we aim to leverage the identified bioactive compounds for applied research not only in drug discovery but also in health foods and cosmetics, thereby promoting research from the agricultural sciences that contributes to a healthier and longer-lived society.
Is more antioxidant protein always better?
While having high levels of antioxidant proteins is generally considered beneficial — reducing susceptibility to alcohol intoxication and cancer, for example — certain cancers have turned this to their advantage, acquiring resistance to anticancer drugs. Antioxidant proteins need to be induced at the right time and in the right organ or location in response to stimuli, and must disappear promptly once the threat has been dealt with.
Evaluation methods
Until now, our analyses have mainly relied on genetically modified mice, but to enable more convenient evaluation we have also introduced experimental systems using small fish. I introduced small fish out of my love for them, and we always welcome collaboration and inquiries from interested parties.
→ For students interested in joining our lab
Research Equipment

Our daily workbench, equipped with protein electrophoresis apparatuses and membrane transfer systems.

A clean bench for aseptic culture of E. coli, essential for plasmid construction used in gene functional analysis.

An incubator for recombinant E. coli culture, set at 37°C. Cultures at lower temperatures (e.g., 20°C) are also carried out as needed.

We maintain four standard PCR machines and two real-time quantitative PCR machines. qPCR detects amplified gene fragments via fluorescent dyes, enabling highly sensitive measurement of gene expression changes.

Our older PCR machines. Each lab member seems to have their own favorite — switching machines can sometimes help when amplification of long fragments proves difficult.

After PCR amplification, agarose gel electrophoresis and staining are used to confirm the product. We also use green light (instead of UV) to minimize DNA damage during visualization — though it is strikingly bright.

Our Milli-Q system further purifies the pure water supplied from the tap (a feature of Saga University's Faculty of Agriculture) to produce molecular biology-grade water. Urban legend has it that drinking it upsets your stomach.

A biosafety cabinet for aseptic culture of mammalian cells, rated for BSL-2 organisms. Our lab is equipped for lentiviral and retroviral gene delivery experiments.

A phase-contrast microscope for observing transparent cultured cells. It has a built-in digital camera so cells can be viewed on a monitor — though I personally prefer looking through the eyepiece.

An inverted fluorescence microscope purchased with a large portion of our research funding. All XYZ stages are fully motorized, enabling high-resolution tiled images of large samples and automated time-lapse photography. Equipped with DAPI, FITC, and TRITC filter sets, it can capture up to four fluorescence channels simultaneously. Pre-programmed coordinates allow fully automated, multi-channel imaging of selected cells at specified time points — the backbone of our visual research.

We primarily use lipofection for gene delivery into cultured cells, but turn to electroporation when high efficiency is required. This pipette-type electroporator is convenient, though the tips are expensive.

A multimode plate reader acquired in a leap of faith after joining Saga University. It supports absorbance, fluorescence (at any wavelength), and luminescence measurement, and can capture bright-field, FITC, and TRITC images of cells in 96-well plates. Invaluable for screening and very popular as shared equipment.

Standard cell culture is performed at 21% O₂ (ambient air), but the body's interior is hypoxic (5–7% O₂). To study physiologically relevant conditions, we use this culture chamber that enables aseptic cell culture at controlled oxygen concentrations between 5% and 21%, with 5% CO₂ and 37°C.

A scanner-type microscope for real-time observation of cells seeded in plates. Particularly useful for monitoring how and when cells die in response to compounds. Installed inside the hypoxia chamber. Word is that a Tadas Lab YouTube channel may be in the works…

Standard CO₂ incubators maintained at 100% humidity, 5% CO₂, and 37°C. The lower unit also allows oxygen concentration control (1.0–20%). Gloves in the lab are used until they wear through, and each pair is labeled with the owner's name — waste not, want not!

One of our many freezers, maintained at −30°C for sample and reagent storage. Each lab member is assigned a dedicated drawer for their experimental samples.

Our ultra-low temperature freezer (−80°C) for long-term storage of RNA, proteins, cultured cells, and tissue samples.